




La déficience en phosphore (P) représente une des carences majeures en éléments minéraux des sols agricoles affectant la production des cultures. Les plantes absorbent le P principalement sous forme d’ions orthophosphates (c’est-à-dire H2PO4- et H2PO4-2). Cependant, la disponibilité des ions orthophosphates pour les racines des plantes est souvent extrêmement faible dans les sols.
La carence en phosphore dans les sols
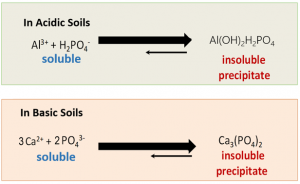
En effet, les sols sont souvent riches en P mais sa phytodisponibilité pour les plantes est affectée par divers processus chimiques et biologiques. Lorsqu’ils sont appliqués au sol, les engrais Phosphatés sont très rapidement immobilisés et transformés en formes P insolubles telles que les phosphates de calcium, dans les sols à pH élevé, oules phosphates d’aluminium ou de fer dans les sols à faible pH, comme le montre la figureci-dessous (Hinsinger, 2001 Plant and Soil. 237 : 173-195 ; Richardson et al., 2009, Crop Pasture Sci.60: 124-143; Penn et Camberato, 201, Agriculture, 9, 120). Les rapports publiés indiquent que seulement moins de 20% de l’engrais P appliqué est absorbé par les plantes tandis que le reste est immobilisé et indisponible pour l’absorption racinaire en raison de la conversion rapide de l’engrais P en formes insolubles.
L’immobilisation microbienne de P est un autre phénomène du sol qui rend l’engrais P indisponible pour l’absorption racinaire, au moins temporairement. Les microorganismes du sol entrent en compétition avec les racines des plantes pour le P disponible et l’immobilisent dans leur biomasse (Richardson et Simpson, 2011, Plant Physiol.156 : 989-996). Il n’est donc pas surprenant que la concentration phyto-disponible (soluble) de phosphore inorganique (Pi) dans la solution du sol soit souvent trop faible (environ 2 µM), ce qui est environ 10000 fois plus faible que les concentrations de Pi existant dans les cellules racinaires (Plaxton et Tran, 2011, Plant Physiol.156 : 1006–1015). Il est intéressant de noter que les cellules racinaires maintiennent des concentrations de Pi très élevées (à des niveaux millimolaires) tandis que la solution de sol contient du Pi à de faibles niveaux micromolaires. Cela suggère que les plantes développent des stratégies racinaires très efficaces pour l’acquisition du phosphore dans les sols (Hinsinger, 2001 Plant and Soil. 237 : 173-195 ; Lynch, 2011, Plant Physiol. 156 : 1041–1049). Ce sujet sera abordé dans un article séparé.
La figure 1 montre que plus de 50% des sols agricoles sont négativement affectés par de faibles quantités de phosphore disponible et présentent un potentiel élevé de rétention / immobilisation du phosphore (Kochian 2012, Nature, 488 : 466-467). La majorité du P existant dans le sol est fermement retenu ou fixé par les particules du sol et n’est pas utile pour l’absorption des racines. L’épuisement du P phyto-disponible par des cultures intensives et l’érosion des sols est un autre problème critique conduisant à une carence en P des sols, en particulier dans les systèmes agricoles à faibles intrants (Tan et al., 2005, J. Sustain. al., 2020, Nature Comm.11: 4546). Selon Alewell et al. (2020), les pertes de P des sols par érosion peuvent dépasser 20 kg ha-1 an-1).
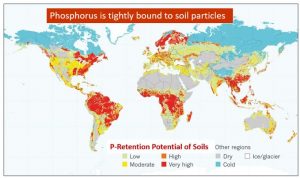
Figure 1. La répartition mondiale des sols différant par le degré de potentiel de rétention du phosphore (immobilisation) (c’est-à-dire que le phosphore est étroitement lié aux particules du sol ou fixé dans la matière organique du sol) (Kochian 2012, Nature, 488 : 466-467).
Dans les sols possédant une capacité de fixation du P très élevée et de faibles quantités de P phyto-disponible, la croissance des plantes est souvent réduite, ce qui entraîne des réductions importantes de la croissance et de la capacité de rendement des plantes cultivées, comme indiqué ci-dessous dans la figure 2 pour le riz cultivé au champ en Tanzanie et aux États-Unis et le maïs aux États-Unis. Les plantes traîtées répondent très positivement à l’application d’engrais P avec une meilleure production de biomasse et un meilleure rendement en grains. Les essais sur le riz menés par le Dr J. Harrell en Louisiane ont montré des augmentations du rendement en grains allant jusqu’à 50%.

Figure 2 : La croissance des plants de riz en Louisiane aux États-Unis (en haut) et en Tanzanie (en bas à gauche) et des plants de maïs aux États-Unis (en bas à droite) avec une fertilisation faible et adéquate en P (voir https://www.ricefarming.com pour les essais de riz par le Dr J.Harrell en Louisiane; africarice.blogspot.com pour une expérience sur le riz en Tanzanie par le Dr De Bauw et Plantsagronomy.k-state.edu pour des expériences sur le maïs)
Les fonctions du phosphore dans les plantes
Le phosphore : un élément de base dans les composantes cellulaires vitales
Le phosphore possède diverses fonctions physiologiques et biochimiques chez les plantes. Il existe plusieurs composés de base dans les systèmes cellulaires qui nécessitent P pour leur structure et leurs fonctions biologiques, notamment l’ADN, l’ARN, l’ATP, le NADPH et les phospholipides.
Le phosphore est indispensable dans le transfert et le stockage d’énergie par l’ATP (adénosine triphosphate) dans les systèmes biologiques et contribue à la synthèse et à la stabilité de l’ADN et de l’ARN. De même, en tant que composant de base des phospholipides membranaires, une nutrition adéquate en P est également essentielle pour l’intégrité structurelle et fonctionnelle des membranes cellulaires (Hawkesford et al., 2012, In: Mineral nutrition of upper plants. Elsevier). Les phospholipides sont le principal composant des membranes cellulaires, nécessaires à la stabilité structurelle et au fonctionnement et au fonctionnement des systèmes de transport membranaire. En cas de carence en P, on peut s’attendre à des altérations structurelles des membranes cellulaires avec des effets néfastes sur le transport des nutriments à travers les membranes des cellules racinaires. Nous présentons et discutons, ci-dessous,quelques fonctions physiologiques critiques du P chez les plantes.
Le rôle du phosphore dans le métabolisme photosynthétique du carbone
Il existe plusieurs rapports traitant du rôle du P dans le métabolisme photosynthétique du carbone (C) et la production de photoassimilats. La majorité des rapports indiquent des fonctions très essentielles du P dans le processus de photosynthèse. En tant que composant structurel de divers phosphates de sucre tels que le ribulose-1,5-bisphosphate (RuBP) et le fructose 6-phosphate, le P affecte grandement la fixation photosynthétique du CO2. En cas de nutrition à faible teneur en P, une altération de la régénération de RuBP est un résultat bien documenté qui diminue l’activité de la Rubisco et par conséquent la fixation du CO2 (Hammond et White, 2008, J Exp Bot 59 : 93–109 ; Warren 2011, Tree Physiology 31, 727–739). La régénération de RuBP dans les chloroplastes est un processus dépendant de l’ATP et, comme discuté ci-dessous, le pool d’ATP est très faible dans les plantes déficientes en P.
Les ions orthophosphates (c’est-à-dire les phosphates inorganiques ; tels que H2PO4-) sont des acteurs importants dans le métabolisme photosynthétique du carbone et sont utilisés comme substrat pour la synthèse d’ATP à travers le processus de photophosphorylation. Par conséquent, une carence en P est souvent associée à une faible concentration d’ions orthophosphates (Pi) dans les chloroplastes, ce qui entraîne une altération de la production d’ATP par l’activité réduite de l’ATP synthase (Figure 3; Carstensen et al., 2018, Plant Physiol 177: 271-284). L’impact négatif d’une faible teneur en P sur la formation d’ATP est hautement dépendante du Pi et se rétablit rapidement après l’application de P aux plantes ayant été carencées en P. Il est donc évident qu’une nutrition adéquate en P est essentielle pour la bonne activité photosynthétique des plantes.
La fixation biologique de l’azote (N2)
Le phosphore s’avère être un nutriment minéral clé responsable d’une fixation biologique efficace du N2 chez les légumineuses. En cas de faible apport en P, le processus de nodulation et le fonctionnement des nodules sont négativement altérés, et la quantité de N fixée est réduite. Les nodules représentent un puits important pour le P. Il a été démontré que la concentration de P des nodules de diverses légumineuses est 3 fois plus élevée que d’autres organes ou partie des plantes (Schulze et Drevon, 2005, J. Exp. Bot. 56 : 1779–84; Qin et al., 2012, Plant Physiol. 159: 1634-1643; Lazai et al., 2017, Agron. J. 109 : 283-290). Qin et al (2012) ont montré que la carence en P altère considérablement la nodulation chez les plants de soja et entraîne une diminution marquée du nombre de nodules et de la taille des nodules (Figure 4). Lorsque les légumineuses souffrent d’une carence en P, une quantité croissante de P est allouée aux nodules.
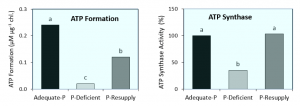
Figure 3 : Les changements dans la formation d’ATP et les activités de l’enzyme ATP synthase dans les chloroplastes des plants d’orge cultivés sous un apport faible et adéquat en P ainsi que dans des plantes déficientes en P après réapprovisionnement en P (Pour plus de détails, voir Carstensen et al., 2018, Plant Physiol 177 : 271-284)
Il existe souvent une relation étroite entre la formation des nodules et la concentration des nodules en P. La forte dépendance de la fixation de N2 à une nutrition suffisante de P est liée au fait que le processus de fixation de N atmosphérique est un processus très exigeant en énergie et nécessite au moins 16 moles d’ATP pour réduire une mole de N2. Par conséquent, l’activité de l’enzyme nitrogénase est également positivement influencée par l’amélioration de l’état nutritionnel P des plantes. Dans les nodules entiers de plants de soja, la concentration d’ATP et la charge énergétique sont significativement réduites en cas de faible apport en P par rapport aux conditions suffisantes en P (Sa et Israel, 1991 ; Plant Physiol.97: 928-935). Ces résultats soulignent l’importance du P dans la performance optimale des nodules et le processus de fixation du N atmosphérique.
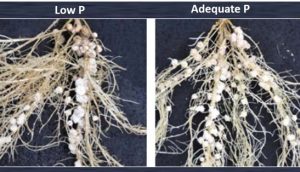
Figure 4 : Les performances de nodulation et de croissance des nodules dans les racines de soja avec une application faible et adéquate de P (Qin et al., 2012, Plant Physiol.. 159: 1634–1643)
Le tallage, l’expansion des feuilles et la croissance reproductrice
L’un des principaux changements structurels en réponse à une faible disponibilité de P dans le milieu de croissance est le processus de tallage. Le tallage et le nombre de talles sont connus pour être des facteurs importants de la plante affectant la capacité de rendement des céréales. En cas de faible apport de P, une réduction ou une suppression sévère du tallage est très courante, ce qui réduit la biomasse et, par conséquent, lerendement (Figure 5 ; Hammond et White, 2008, J Exp Bot 59 : 93–109;). À l’instar du tallage, l’expansion des feuilles est également très sensible à la carence en P, et une altération de l’expansion des feuilles en réponse à un faible apport en P a été suggérée comme le premier changement chez les plantes déficientes en P (Radin et Eidenbock, 1984, Plant Physiol 75 : 372 à 377 ; Lynch et al., 1991 ; Crop Sei. 31 : 380 à 387). Une diminution de la surface foliaire diminuera l’interception de la lumière et la production de biomasse. La réduction de l’expansion foliaire en cas de faible apport de P a été attribuée à un transport réduit de l’eau du milieu de croissance vers les feuilles et donc à une faible pression de turgescence nécessaire à l’expansion.

Figure 5 : Des plants de blé cultivés sur un sol déficient en P avec une fertilisation faible, moyenne et adéquate en P (photo A. Yazici et I. Cakmak)
La carence en phosphore a également un impact négatif évident sur la croissance reproductive des plantes. Le retard du développement reproducteur et le retard de la maturation sont des problèmes courants chez les plantes à faible apport en phosphore. Cependant, selon Nord et Lynch (2008, Plant, Cell and Environ 31: 1432–1441), le retard observé de la floraison et de la maturité en cas de carence en P pourrait être un retard utile pour les plantes: il peut permettre aux plantes d’avoir plus de temps pour l’utilisation et la remobilisation du P des tissus végétatifs.
Les symptômes de carence en phosphore
Les symptômes de carence en phosphore commencent généralement sur les feuilles plus âgées en raison de la mobilité élevée du phloème du phosphore. Habituellement, les symptômes de carence visuelle en P se développent dans les feuilles contenant moins de 0,2 pour cent de P sur la base du poids sec. Le développement d’une couleur vert foncé et bleuâtre sur les feuilles est un symptôme caractéristique d’une carence en P chez les plantes (figure 6). Lorsque la carence en P est avancée, la chlorose des feuilles et la nécrose du bout des feuilles peuvent également être visibles. Dans des conditions de carence en P, la croissance des pousses est connue pour être plus affectée que la croissance des racines, très probablement, parce que les plantes carencées en P allouent des quantités plus élevées de photoassimilats dans les racines pour une meilleure formation et adaptation des racines. Cette question sera abordée dans un autre article.

Figure 6 : Jeunes plants de maïs cultivés dans un milieu avec une application faible et adéquate de P (photo A. Yazici et I. Cakmak)
Conclusion
Aujourd’hui, la plupart des sols agricoles ont des problèmes de carence en P limitant la productivité des cultures, tandis que dans certaines régions, les excédents en P sont courants. Le problème de carence en P du sol est encore aggravé par l’épuisement du P du sol en raison de la culture continue et de l’érosion du sol. Le suivi et la mesure de l’état nutritionnel des plantes par l’analyse du sol et des tissus végétaux est une pratique agronomique importante pour réduire les altérations de la croissance des plantes liées à une carence en P. Cependant, comme les analyses de sol ne sont généralement pas forcément utiles, les résultats de l’analyse des tissus foliaires pour P devraient être intégrés aux tests de P du sol. Il est suggéré que l’analyse des plantes pour le statut P devrait être réalisée plutôt au début de la croissance. On sait que le niveau de disponibilité de P pendant les premiers stades de croissance est très critique et a un effet décisif sur le potentiel de rendement des plantes cultivées (Grant et al., 2001, Can. J. Plant Sci. 81 : 211–224) .